
















This article was written by Serge, MSc. Plant Biologist and Environmental Scientist with a BSc in Plant Biology and an MSc in Environmental Biology and Biogeochemistry. My research focused on climate change effects on boreal forest ecosystems. I write from field experience, not just literature.

I want to be upfront about something before I explain what I found. I measured growth. I measured soil respiration. I did not measure the biochemistry happening inside the leaves, the antioxidant enzyme activity, the phenolic compound concentrations, the stomatal conductance responses. Those measurements would have told me exactly why gt14 and gt15 behaved differently under the same treatments. I did not have that data.
What I had was the pattern. And the pattern was genuinely interesting.
Two genotypes of the same species, silver birch, growing side by side in the same soil, exposed to identical temperature and ozone treatments. One responded one way. The other responded differently. Sometimes opposite. That genotype-dependent pattern ran through almost every measured parameter and it told me something real about how genetic variation shapes plant responses to environmental stress, even without the biochemical explanation for why.
This article is my interpretation of what I observed, what I think was probably happening, and what questions the data left unanswered.
The Experiment
The field site was an open-air exposure system using eight circular plots, each ten metres in diameter. Four treatment combinations: control with ambient air and temperature, elevated temperature alone, elevated ozone alone, and combined elevated temperature plus ozone.
Temperature elevation was achieved with infrared heaters above the canopy maintaining a plus 0.9 degrees Celsius increase over ambient throughout the growing season. Not dramatic warming. Deliberately modest, designed to reflect near-future climate projections for high-latitude boreal regions rather than extreme scenarios.

Ozone fumigation delivered concentrations of approximately 33.4 parts per billion in treatment plots compared to 24.2 parts per billion in ambient plots. The ozone was applied 14 hours per day, seven days per week. The AOT40 accumulated ozone exposure value reached 4.4 parts per million per hour in treatment plots versus 0.1 in controls.
Two silver birch genotypes, gt14 and gt15, grew in separate sections of each subplot separated by root exclusion cloth so their root systems could not intermingle. This was important because soil respiration measurements needed to reflect each genotype’s own root and associated microbial activity rather than a mixed signal.
I measured stem height and diameter every three weeks from June through September. I measured leaf count and leaf area on marked branches. I measured soil CO₂ efflux monthly using a LICOR chamber at two centimetre depth alongside soil temperature and moisture.

What Temperature Did
Warming enhanced stem height significantly. The effect peaked in mid-July at roughly nine percent taller than control trees, settling to around three to four percent by late August as the season progressed. Both genotypes showed this response. Temperature clearly benefited stem extension growth.
Soil respiration increased substantially under warming. Gt15 showed a 36 percent increase in soil CO₂ efflux under elevated temperature. Gt14 showed 24 percent. Both significant. Both in the same direction.
My interpretation of the soil respiration increase: warming stimulated root metabolic activity and the microbial community feeding on root exudates simultaneously. The CO₂ coming out of the soil represents both root respiration and microbial decomposition activity, and both respond to temperature. I covered this mechanism in detail when discussing what I measured in that experiment in the context of soil food web biology elsewhere on this site.
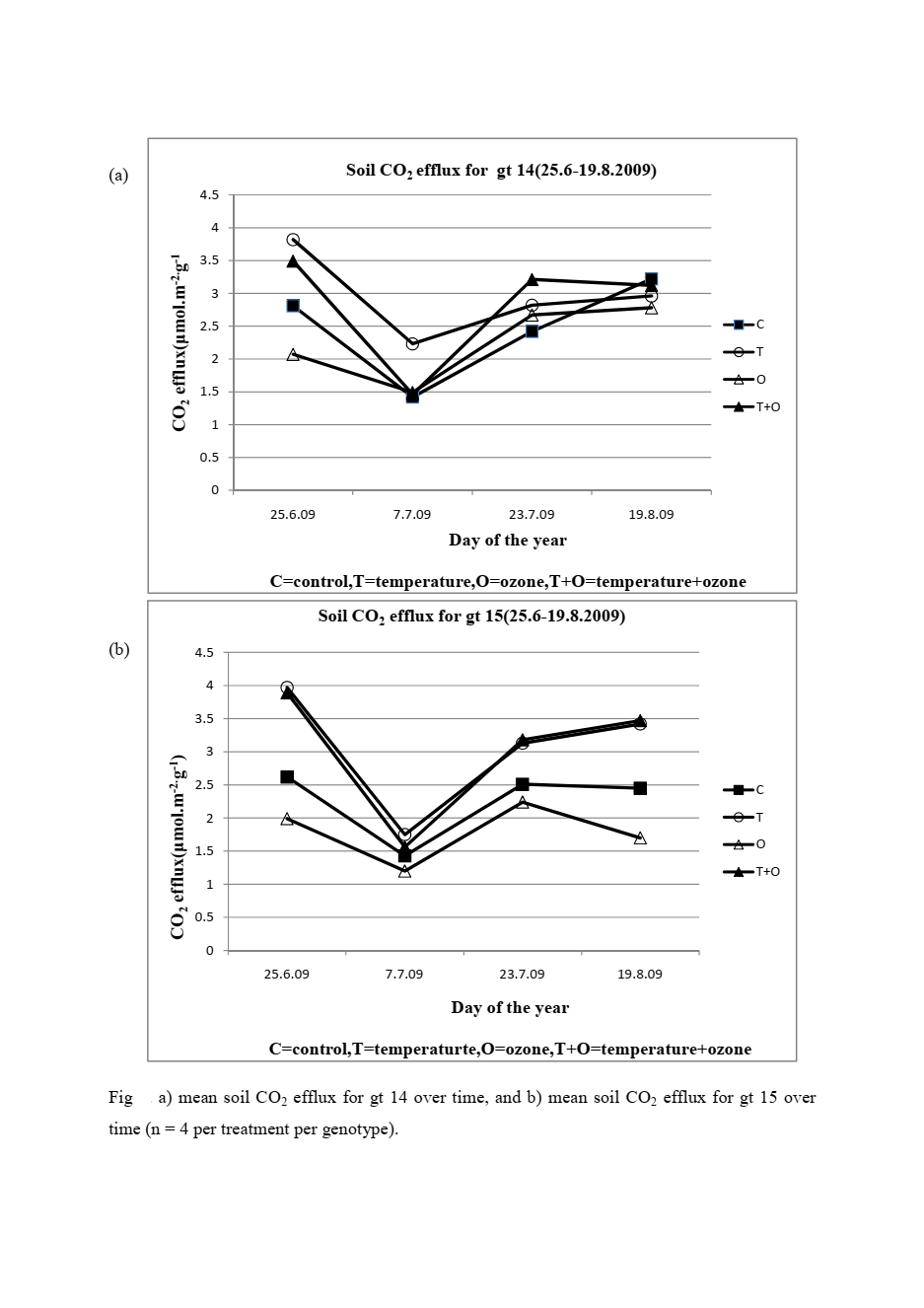
What I found interesting about the difference between genotypes in soil respiration response is that gt15 responded more strongly. My best guess is that gt15 had a larger root system by this point in the season, giving it more root surface area contributing to respiration. I did not measure root biomass so I cannot confirm this. But gt15 was consistently the more vigorous genotype across most above-ground growth parameters, and more vigorous above-ground growth typically correlates with more extensive below-ground investment. It would fit.
The Leaf Area Finding That Surprised Me
This is where the results got genuinely interesting and where I wish I had measured more.
Temperature increased total leaf area in gt14. That makes intuitive sense. Warmer conditions, more growth, more leaf area for photosynthesis.
Temperature decreased total leaf area in gt15.
Same treatment. Opposite direction. Both statistically significant.
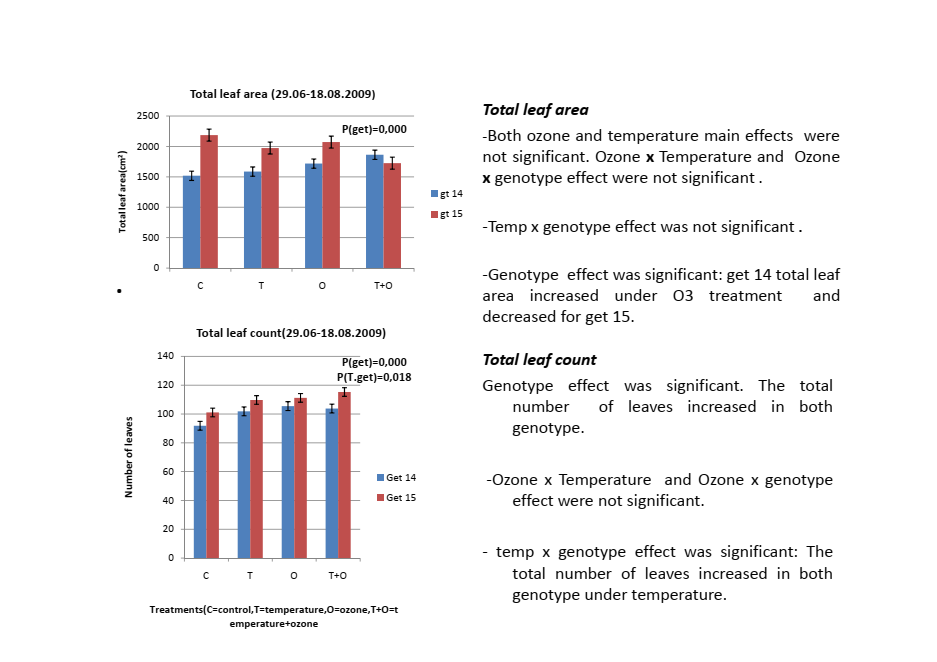
I remember looking at these results and thinking, wait. The more vigorous genotype, the one with more leaves and thicker stems, responded to warming by producing smaller leaves? That seemed counterintuitive.
The most plausible explanation I can offer, and I want to be clear this is my interpretation rather than something I measured, is that gt15 may have been investing more in leaf number at the expense of individual leaf size under warming. Gt15 produced eight to ten percent more leaves than gt14 under temperature treatment. If total carbon allocation to leaves remained similar between the treatments, more leaves of smaller size is one possible outcome.
There is also a mechanism suggested in the literature involving increased photorespiration under warming, where the ratio of oxygenation to carboxylation reactions by Rubisco shifts at higher temperatures, reducing net carbon gain per leaf. If gt15 has different Rubisco kinetics or thermal acclimation capacity than gt14 this could explain why warming reduced its leaf area. But I did not measure any of that. I am speculating based on what I know from plant biochemistry training and what the data pattern suggests.
What I can say with confidence is that the two genotypes acclimate to warming differently at the leaf level. That is a real finding regardless of the biochemical explanation.
What Ozone Did, and What It Did Not Do
Here is the answer: in a single growing season at these concentrations, ozone did not dramatically affect stem height or leaf growth in either genotype.
I expected ozone to reduce growth more than it did. Previous studies had shown significant growth reductions in silver birch under elevated ozone. Our AOT40 exposure was not trivial at 4.4 parts per million per hour. But the above-ground growth effects during this first season were modest.
The one ozone effect that was statistically clear was a late-season reduction in stem diameter in gt14 under ozone alone. Not in gt15. Not under the combined temperature plus ozone treatment. Just gt14 under ozone alone, appearing in the measurements toward the end of August.
Why gt14 and not gt15? I have thought about this. My best guess involves two possible mechanisms, and I genuinely do not know which is more important.
First, gt15 consistently showed more vigorous growth throughout the experiment. More vigorous plants often have higher antioxidant capacity, more resources available to repair oxidative damage, and potentially greater capacity to produce the protective volatile organic compounds that silver birch releases when stressed. I believe gt15 may simply have been better equipped to cope with the oxidative stress ozone generates inside leaf cells. But I did not measure antioxidant enzyme activity or VOC emissions in my experiment so I cannot confirm this.
Second, the ozone effects on stem diameter appeared late in the season. By that point gt14 and gt15 had been growing under different conditions for months and their physiological states were diverging. Whether gt14 was more depleted in carbon reserves by late season, less able to sustain cambial growth under additional oxidative stress from ozone, I cannot say for certain. It is plausible.
What I am confident about is that genotype matters. The same ozone exposure at the same concentration produced different outcomes in two genotypes of the same species. That has real implications for thinking about which forest tree populations might persist and which might struggle under increasing tropospheric ozone as climate change continues.
The Most Interesting Finding: Warming Protected Against Ozone
This was the result I found most conceptually interesting and that connects most directly to broader questions about how climate change and air pollution interact.
The ozone-induced reduction in gt14 stem diameter I described above appeared under ozone alone. Under the combined temperature plus ozone treatment it disappeared. Warming appeared to counteract the ozone effect on stem diameter growth.
There are at least three plausible mechanisms for this and I want to be honest that I do not know which operated in our experiment.
Warming can reduce stomatal conductance in some species, reducing the amount of ozone that enters leaf tissue per unit time. Less ozone uptake means less reactive oxygen species generation and less damage. This could explain why combined warming plus ozone produced less diameter reduction than ozone alone.
Warming increases isoprene and other volatile organic compound emissions from many tree species. These compounds have antioxidant properties inside leaf cells and can quench reactive oxygen species before they cause membrane damage. Silver birch is not a high isoprene emitter under normal conditions but warming does increase its VOC emissions. I think this mechanism is plausible for our results but again I did not measure VOC emissions in my experiment.
Enhanced carbon assimilation under warming could simply provide more substrate for repair processes, allowing the tree to maintain cambial growth despite some ozone damage that it could not sustain under ambient temperature.
All three mechanisms could operate simultaneously. The data cannot distinguish between them. What the data shows is the outcome: warming mitigated the ozone effect on stem diameter. The why remains an open question from my experiment.
What I Wish I Had Measured
Being honest about the limits of what I measured is part of being a scientist rather than just a data reporter.
I wish I had measured stomatal conductance. Knowing how gt14 and gt15 regulated their stomata differently under the four treatments would have told me directly how much ozone each genotype was actually taking up. That single measurement would have explained much of the genotype-specific pattern I observed.
I wish I had measured leaf phenolic content and antioxidant enzyme activity. If gt15 was producing more protective compounds than gt14 under ozone stress that would explain why it tolerated the ozone treatment better for most parameters while gt14 showed the late-season diameter reduction.
I wish the experiment had continued for multiple growing seasons. The ozone effects I measured were mostly minor in a single season. The literature consistently shows that ozone effects accumulate. Whether the genotype-specific patterns I observed in year one would have amplified, reversed, or remained stable over two or three seasons is genuinely unknown and would have been valuable to know.
These are not criticisms of the experimental design. Single-season field experiments with four treatment combinations already generate enough data complexity to fill a thesis. But the questions that remain unanswered are sometimes more interesting than the ones the data answered.
What This Means Beyond the Birch Plots
The genotype-dependent responses I measured in silver birch have implications that extend beyond this one experiment.
Forest tree populations contain genetic variation in stress tolerance. Some individuals within a species are better equipped than others to handle warming, ozone, drought, and their combinations. As climate change and tropospheric ozone concentrations both increase, the composition of forest populations will shift toward individuals with the genetic traits that confer stress tolerance.
Understanding which traits those are, whether antioxidant capacity, stomatal regulation, VOC emission rates, or carbon allocation strategy, would help predict how forest productivity and carbon cycling will change. My experiment contributed one data point to that larger question: gt14 and gt15 respond differently, genotype matters, and the direction of the effect depends on which stressor you are measuring and when in the season you look.
That is a summary of what I found.
FAQs
What did your field experiment measure?
Stem height and diameter, leaf count and leaf area, and soil CO₂ efflux across two silver birch genotypes under four treatment combinations: control, elevated temperature, elevated ozone, and combined temperature plus ozone. The experiment ran through one full growing season measuring responses every three weeks for growth parameters and monthly for soil respiration.
Why did the two genotypes respond differently to ozone?
I cannot give a definitive answer because I did not measure the biochemical responses. My best interpretation is that gt15 may have had higher antioxidant capacity, more vigorous growth providing more carbon for repair processes, or different stomatal regulation that reduced ozone uptake compared to gt14. The late-season reduction in gt14 stem diameter under ozone alone suggests gt14 was more vulnerable to ozone oxidative stress at that growth stage.
Why did warming reduce the ozone damage?
Three plausible mechanisms: reduced stomatal conductance under warming limiting ozone uptake, increased protective VOC emissions under warming quenching reactive oxygen species inside leaf cells, and enhanced carbon assimilation under warming providing more substrate for cellular repair. My experiment cannot distinguish between these mechanisms. The outcome, warming mitigated ozone damage to stem diameter, is clear in the data. The why requires measurements I did not take.
Were the ozone effects large?
In a single growing season at these concentrations, the effects on most measured parameters were modest. The most consistent finding was the late-season reduction in gt14 stem diameter under ozone alone. The literature suggests ozone effects accumulate over multiple seasons, so single-season experiments likely underestimate long-term growth impacts.
What would you measure if you repeated the experiment?
Stomatal conductance to quantify actual ozone uptake by each genotype. Leaf phenolic and antioxidant enzyme activity to understand why the genotypes differed in ozone tolerance. Multiple growing seasons to assess cumulative ozone effects. VOC emission rates to test whether warming-induced VOC increases contributed to the protective effect against ozone damage.
Does this have implications for forest management?
Yes. Genotypic variation in stress tolerance exists within silver birch populations and likely within most forest tree species. As warming and tropospheric ozone both increase under climate change scenarios, forest composition will shift toward individuals with traits conferring stress tolerance. Understanding which genotypes are more tolerant and why helps predict forest productivity and carbon cycling under future conditions.